compiled by Brian Tomasik
First published: 4 Jan. 2017; last update: 18 Nov. 2017
Summary
This page collects information on the "cognitive" abilities of unicellular organisms. This topic is important because most of the heterotrophic metabolism on Earth is done by microbes rather than animals.
See also: "Microbial intelligence"
Contents
Numerosity of microbes
Why is it important to assess the sentience of unicellular organisms? One reason is just that the case of unicellular creatures provides a useful testing ground when evaluating one's theories of consciousness. However, the issue also has potentially practical importance, because unicellular organisms are the dominant form of heterotrophic life on Earth in terms of metabolism and biomass. Thus, if unicellular organisms do warrant even tiny amounts of moral concern, the implications for our ethical behavior could be immense.
Smil (2003) estimates that almost all of the Earth's heterotrophic biomass takes the form of prokaryotes.
Brady (1974) reports that "80-90 percent of the total soil metabolism is due to the microflora" (p. 113)a, which are "microorganisms, including algae, fungi, and bacteria that live in or on another living organism or in a particular habitat". Meanwhile, Petersen and Luxton (1982) report regarding animals: "The soil fauna appears generally to be responsible for less than about 5% of total decomposer respiration" (p. 288).
General discussion
Bray (2009), p. 18:
single cells are aware of their surroundings. They detect chemical flavors, mechanical vibrations, visual stimuli, electric fields, and gravity. They respond by moving or by changing their shape or internal state, selectively, in a discerning manner.
Consider a simple organism—say a planarian or an amoeba—moving nonrandomly across the bottom of a laboratory dish, always heading to the nutrient-rich end of the dish, or away from the toxic end. This organism is seeking the good, or shunning the bad—its own good and bad, not those of some human artifact-user. Seeking one’s own good is a fundamental feature of any rational agent, but are these simple organisms seeking or just ‘seeking’? We don’t need to answer that question. The organism is a predictable intentional system in either case.
In light of the information discussed in this piece, as well as the sometimes impressive cognitive abilities of plants, my general sense is that—whether a species has a conventional nervous system or not—if you apply hundreds of millions of years of evolutionary optimization power, the resulting life forms will likely have a lot of intelligent tricks up their sleeves.
Bacteria
Bacteria exhibit the basic properties of living organisms, including growth and reproduction, environmental sensing, and responsive behavior.
Bryce Huebner notes:
I'm in the process of writing a tiny bit of a paper right now about bacterial behavior. It's in the context of a broader discussion of group behavior, and I realized that it was little more than bad ideology that leads philosophers to focus on complex prokaryotes in thinking about adaptive forms of group behavior. One of the things that keeps striking me as I read through the relevant literature is that the behavior of these tiny machines is really complex and highly adaptive. Their preferences might be simple, and they're probably grounded in little more than an impulse to pursue nutrients and avoid toxins, but bacteria seem to pass the William James test for mentality (fixed ends, with varying means). This is leading me to think that the whole idea of mindedness is poorly conceived, or that bacteria are minded...
This article quotes professor Jeffrey Lichtman:
Minds drum up the feeling of pain to keep the body safe, he continued, but the animal kingdom is full of self-preserving behaviors. Lichtman even suspected “that the first living animals that could affect their position in the world by moving very quickly developed ways to avoid putting themselves at risk, and that is equivalent to pain.” He and Dennett agreed that even microbes have incentives to sense their surroundings and avoid harm analogously to how humans avoid pain.
Bray (2009), pp. 5-7:
In 1854 the German biologist Wolfgang Pfeffer showed that if he introduced a capillary pipette filled with nutrient mixtures such as yeast or meat extract into a solution containing swimming bacteria, the bacteria would collect around the pipette and eventually enter into its tip. Capillaries filled with acid, alkali, or alcohol had the opposite effect, causing the bacteria to swim away. Other investigators at the time observed bacteria responding to light, temperature, or the concentrations of salts. [...]
Escherichia coli can detect something like fifty distinct chemicals. The list includes sugars and amino acids that act as attractants (the bacterium swims toward them) and a motley mixture of heavy metals, acids, and toxic substances that are repellents. E. coli's sensitivity is legendary. Even the slightest whiff of the attractant amino acid aspartate (a concentration of less than one part in ten million) is enough to change its swimming. [...]
to pursue any direction for more than a second or so, bacteria have to continually reassess their situation. How do they do this? The answer is that they have a sort of short-term memory that tells them whether conditions are better at this instant of time than a few seconds ago. By "better" I mean richer in food molecules, more suitable in acidity and salt concentration, closer to an optimum temperature, and so on. If on average conditions have improved, or at least are not any worse, then the bacteria will continue to swim in the same direction. But if conditions are deteriorating, then the bacteria tumble; they swim off in a new direction, selected more or less at random. The repeated execution of this pragmatic routine carries them over long distances and complicated terrains toward favorable locations.

This video says: "It's fair to say that E. coli has a short-term memory, but it's only about 4 seconds long. So what the cell does is it counts the things of interest in its environment over a second, looks up what it got for the previous 3 seconds, and then responds to the difference."
Freeman (2002) says (p. 274) regarding bacteria:
Although some species specialize by using just one type of food, the vast majority of bacteria are able to switch among several distinct sources of carbon and energy depending on which food items are available in the environment. [...]
[Intestinal E. coli] cells are capable of using a wide array of sugars to supply the carbon and energy they need. As your diet changes from day to day, the availability of different sugars in your intestine varies. Precise control of gene expression gives E. coli cells the ability to respond to these changes in its environment and use the different sugars.
Freeman (2002) also describes an experiment by Jacques Monod in the 1950s to test the hypothesis "that E. coli might actually prefer glucose to lactose as a food source" (p. 277). The experimental results were consistent with this hypothesis.
Fernando et al. (2008): "Although adaptive sensitization and habituation (non-associative learning) have been demonstrated in bacteria (Yi et al. 2000), associative learning within a single lifetime has not. An example of sensitization is the autocatalytic upregulation of phoA and phoB after prior phosphate limitation, resulting in a stronger response to subsequent phosphate limitation (Hoffer et al. 2001)."
Habituation and sensitization have both been demonstrated in bacterial [chemotaxis] (Koshland et al., 1982; Stock, 1999; Porter et al., 2011)—a discovery that “gave some neurophysiologists apoplexy, because they believed that a nervous system” was required (Taylor, 2004, p. 3761). [...]
Researchers in two of the studies described here (Tagkopoulos et al., 2008; Mitchell et al., 2009) claim their discoveries are demonstrations of associative learning. However, conditioning in these cases is clearly epigenetic. Whether this will count as ‘genuine conditioning’ remains an open question.
Groups of bacteria appear to coordinate via quorum sensing. Even if individual bacteria don't matter much, it's plausible that the entire colony matters more.
Protists
This video shows the behavior of many types of protists.
Applewhite and Morowitz (1967), pp. 332-33:
While protozoa lack the neurons of the metazoa, they do possess a fibrillar network that presumably may carry out the same functions (Kudo, 1966). Electron microscopy has been done on many of them (Pitelka, 1963), and they exhibit a wide variety of behaviors (Jennings, 1962, and Warden et at., 1940) that could be used to elicit learning. [...]
There have been many learning experiments with protozoa (McConnell, 1966, and Thorpe, 1963), but true learning in them has been questioned (McConnell, 1966).
Amoeba
Bichai et al. (2008), p. 510:
Free-living amoebae generally have 2 stages of development: the trophozoite and the cyst (Greub and Raoult 2004). The trophozoite is the active metabolic stage, feeding on bacteria and multiplying by binary fission. Hostile pH conditions, osmotic pressure, temperature, or even unfulfilled nutritional needs of the amoeba can cause its encystment (Greub and Raoult 2004).
In other words, amoebae can detect environmental conditions and respond adaptively to them.
Bray (2009), pp. 11-12:
A single cell of Amoeba proteus is a tiny speck of cytoplasm less than half a millimeter in length. Its amorphous, highly changeable form features fingerlike projections—pseudopodia—from a central mass. The cell lives on the bottoms of streams and ponds and typically crawls over the surfaces of pebbles and vegetation by extending and retracting its pseudopodia. When it encounters obstacles in its path, regions in contact with the obstacle become quiescent while those farther away move more actively. Consequently the cell flows in a new direction. Other modes of movement are also seen. For example, a hungry amoeba often adopts a more rapid form of progression termed loping, alternately extending and contracting its body like an inchworm. Another state I like to think of as skydiving occurs when an amoeba is freely suspended in water. The cell now adopts a starlike form with long pseudopodia extending in all directions. It seems to be searching for a suitable surface. Indeed, no sooner does one pseudopodium make an effective contact than currents of cytoplasm flow in that direction. Other pseudopodia retract; the cell settles onto the surface and resumes its normal mode of creeping.
[...] Herbert Jennings, a naturalist working at the Zoological Laboratory of the University of Michigan in the late nineteenth century, observed spherical cysts, or spores, of Euglena (a ciliated protozoan) being used as food [by Amoeba proteus]. The amoeba in his cultures attempted to engulf the cysts by extending lateral pseudopodia and a thin sheet of cytoplasm. If the attempt was successful, then the cyst was enclosed in a cup-shaped depression, entered the cell in a parody of swallowing, and was eventually digested. The cysts, however, were smooth and rolled easily when touched. Jennings recorded an amoeba that pursued a rolling cyst in a circular path for fifteen minutes, making repeated attempts at capture, until the cyst was eventually lost. "This behavior," Jennings remarked "... makes a striking impression on the observer who sees it for the first time. The Amoeba conducts itself in its efforts to obtain food in much the same way as animals far higher in the scale."
Rather surprisingly, Amoeba proteus is also able to capture actively swimming organisms such as the ciliated Paramecium despite its lumbering gait. It seems to be able to detect swimming paramecia at some distance and may pursue them over long distances. Once contact is made, the amoeba initiates a slow encircling action and attempts to trap the struggling prey. If encounters with swimming prey are sufficiently frequent, the amoeba anticipates a capture by forming a cup-shaped depression on its surface like a baseball mitt.
Bray (2009), p. 18:
An amoeba can immediately tell a Euglena cyst from a grain of sand of the same size and will devour the former while rejecting the latter.
Paramecium
While Stuart Hameroff's views on consciousness are dismissed by most scientists, Hameroff does make an important observation about the abilities of unicellular organisms that lack nervous systems:
A single celled paramecium swims around. It finds food. It finds a mate. It has sex. It can learn. It can avoid predators. It can avoid obstacles. It's one cell. It does it with microtubules.
Fernando et al. (2008):
Hennessey (1979) showed that the single-celled ciliate Paramecium caudatum might possibly be capable of being classically conditioned. A paramecium was successfully trained to exhibit an avoidance response to a conditioned vibration stimulus, using an electric shock as the unconditioned stimulus. This response persisted over the entire lifetime of the paramecium, although no inheritance of this response was studied. Associative sensitization and pseudo-conditioning were not ruled out in the experiment, the repetition of which we would welcome.
Stentor
Wikipedia says that Stentor "are among the biggest known extant unicellular organisms."
Video: "Stentor hunting for food".
Bray (2009), pp. 14-17:
Unexpectedly, the most intricate behaviors so far recorded in unicellular animals have been found in organisms attached to a surface rather than swimming free. The trumpet-shaped Stentor spends most of its time attached to water plants or debris by an irregularly shaped foot embedded in a transparent tube of mucus. Its slender body and flattened, discoid mouth cavity are lined with fine cilia. Beating of these cilia sets up continual currents of fluid that carry particles of suitable size into the mouth cavity. The animal then decides whether they are edible or not. Particles of food are retained and ingested, whereas indigestible particles are ejected. The organism can also turn or sway on the end of its long stalk, thereby pointing its mouth cavity in selected directions.
The lynx-eyed Herbert Jennings gave a detailed account of the response of Stentor roeselii to experimental stimulation. A fine current of water squirted at the open disk caused the cell to contract in a flash into its tube. After about half a minute the animal extended again and the cilia resumed their activity. A second application of the jet of water identical to the first, however, was then ignored, and the animal continued with its feeding activities. Jennings observed a similar sequence of a "surprise" response followed by rapid acclimatization with a whole variety of stimuli, such as a small jarring of the animal's container. Once again the organism displayed a simple form of memory.
Jennings found, however, that a potentially injurious stimulus, such as a cloud of particles of the red dye carmine introduced to the feeding disk, evoked a richer set of actions. The stentor at first makes no response, taking the obnoxious particles of carmine into the pouch and into the mouth. After a short time, though, it turns to one side by bending its long stalk (always in the same direction) as if to move its mouth out of the path of the cloud of noxious particles. This may be repeated: the animal twists on its stalk two or three times about its long axis. If the repeated turning to one side does not relieve the irritation, a second strategy is tried. Beating of the ciliary hairs on the body is suddenly reversed in direction, thereby causing particles in the disk and in the pouch to be ejected. The reversal lasts but an instant before the usual current is resumed. If the irritant particles continue to arrive, the reversal is repeated two or three times in rapid succession.
If stentor still does not get rid of the stimulation in either of the ways just described, it contracts into its tube. In this way, of course, it escapes the offending cloud, but at the expense of losing all opportunity to obtain food. The animal will now remain in the tube about half a minute before again cautiously extending. When the body has reached about two-thirds its original length, the cilia around the disk-shaped mouth begin to unfold. They recommence their beating, allowing the animal to recommence feeding.
What happens now? Suppose that the water currents again carry noxious carmine grains into the feeding disk. The stimulus and all the external conditions are the same, so will the stentor go through the same routine—bend to one side, reverse its cilia? No. The animal has become changed by its experience and tries a new strategy. This time, no sooner do the carmine particles reach its disk than the animal contracts fully into its tube. The sequence may be repeated many times, the animal cautiously extending after a while and, if it still finds particles, contracting again. Each time, it stays a little longer in the tube and seems to be more reluctant to emerge. Finally, it contracts violently and repeatedly while still enclosed in its tube, so violently that the foot attachment breaks and the animal is set free. Stentor leaves its tube and swims away, avoiding if necessary the cloud of carmine and other obstacles, and sets sail for a new home.
In due course, the minute creature will settle onto the bottom of its pond or container. It then behaves in a peculiar way. The partially unfolded disk (the part previously used for feeding) is now used as a means of locomotion. Stentor now creeps rapidly over the terrain, following all the irregularities of the surface rapidly and seeming to explore it thoroughly. This may last for sometime, the animal often quitting one site and swimming on to another, exploring heaps of debris and surfaces of solids in its path.
After ~10-20 minutes, the stentor decides on a location, builds a new mucus tube for itself to live in, and attaches to its new home (Bray 2009, p. 17).
Bray (2009), p. 18:
Most microorganisms display what in higher animals is termed attention. A stentor or amoeba dislodged from a surface actively seeks a new site of attachment. While engaged in this search it ignores other stimuli such as changes in temperature or chemical signals that produce an immediate reaction in a free-living individual.
Unicellular plants
Chlamydomonas
Bray (2009), p. 18:
Almost all single-celled organisms respond to light. Ciliates such as paramecia move to weak light but are repelled by strong light. The single-celled alga Chlamydomonas has a so-called eyespot containing pigment molecules that enable it to detect the direction of incoming light.
Gene regulation
"A stimulus from the environment causes the biological entity to respond." Ordinarily when we hear this statement, we picture an animal transmitting information via its nervous system in order to move its body. Perhaps we might also picture plants responding to environmental stimuli such as the direction of the sun or alarm signals from neighboring plants. However, this statement can also apply to basically every individual living cell, including unicellular organisms. A prominent example of why is that living cells carry out regulation of gene expression, i.e.:
a wide range of mechanisms that are used by cells to increase or decrease the production of specific gene products (protein or RNA)[...]. Sophisticated programs of gene expression are widely observed in biology, for example to trigger developmental pathways, respond to environmental stimuli, or adapt to new food sources.
A famous example is regulation of the lac operon in many enteric bacteria.
Gene regulation is not ordinarily thought of as a form of intelligence, but in some sense, the quasi-digital "computing" machinery of cells that allows them to turn on and off a variety of genes in complex ways is arguably as impressive as what human-made computers can do. So if we think human-made computers have some degree of sentience, perhaps we should think that gene regulation in cells does as well.
Wikipedia's article on "Bacterial stress response":
Bacteria can survive under diverse environmental conditions and in order to overcome these adverse and changing conditions, bacteria must sense the changes and mount appropriate responses in gene expression and protein activity. The stress response in bacteria involves a complex network of elements that acts against the external stimulus. Bacteria can react simultaneously to a variety of stresses and the various stress response systems interact (cross-talk) with each other. A complex network of global regulatory systems leads to a coordinated and effective response.
Boor (2006) discusses "how single-celled organisms respond to environmental stresses." One key concept is sigma factors: "In bacteria, alterations in gene expression are often controlled at the transcriptional level through changes in associations between the catalytic core of RNA polymerase and the different sigma factors present in a bacterial cell [...]. Associations between different alternative sigma factors and core RNA polymerase essentially reprogram the ability of the RNA polymerase holoenzyme to recognize different promoter sequences and express entirely new sets of target genes." The "regulon" is "the set of genes controlled by a single sigma factor". σB is one type of sigma factor found in Bacillus subtilis and other bacteria. Boor (2006) says: "The B. subtilis σB-dependent general stress regulon is large: over 200 genes are expressed following bacterial exposure to heat, acid, ethanol, salt stress, entry into stationary phase, or starvation for glucose, oxygen, or phosphate".
Boor (2006) notes that there can be "genes that may appear to be unrelated in function, but that must be coordinately regulated to enable an organism to survive and respond appropriately under rapidly changing environmental conditions". I picture sigma factors as like broadcast signals that cause updates to potentially lots of different subcomponents of the cell at once. Sigma factors might be compared with neural broadcasts that cause updates to lots of brain processes at once in the global workspace theory of consciousness.
No consciousness without representation?
One idea I sometimes hear is that consciousness requires mental representations, where "A mental representation [...] is a hypothetical internal cognitive symbol that represents external reality". Do unicellular organisms have mental representations?
In a very crude sense, they do. For example, a bacterium's sigma factors "represent" environmental conditions and facilitate appropriate responses.
However, another line of argument is to question the necessity of internal mental representations. In a famous paper, Brooks (1991) wrote (p. 140):
When we examine very simple level intelligence we find that explicit representations and models of the world simply get in the way. It turns out to be better to use the world as its own model.
A cognitive representation is a simplistic model of complex real-world phenomena. But why can't the real-world phenomena themselves be an ethically relevant "mental representation", where this representation exists in the "extended mind" of an organism rather than within its own body? Indeed, the real world is a vastly more sophisticated "model" than what's present in an organism's internal cognition.
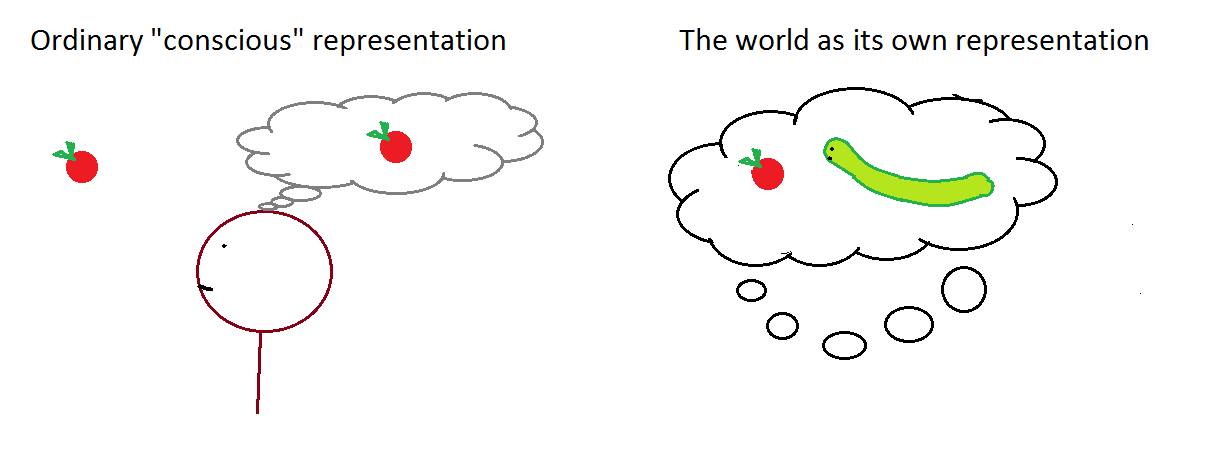{kind=link}
The usual picture of consciousness looks like this, where "-->" means "gives rise to", and words in green are "conscious":
world --> complex mental representations in the organism's mind --> thoughts/actions/etc.
But why can't we imagine the world as a complex "mental" representation of itself, such that thoughts and actions taken directly in response to it can be seen as "utilizing complex representations"?
world (which is a complex representation of itself) --> thoughts/actions/etc.
Organisms who interact with the world can be said to be "using" that "cognitive" representation. For example, a paramecium that eats bacteria uses a complex "mental representation" of the world (namely, the world itself) when seeking out and devouring its food.
The objection might be raised that this is "cheating" and that you need to have an intermediate layer of complex representations in the organism's own cognitive system. I would ask: "Why? What's so special about internal representations?" The best response I can imagine would be along the following lines. As species become more intelligent, they increasingly move representations from the external world into their internal cognitive systems because this allows for more sophisticated, high-level concepts. For example, a unicellular organism can respond to various signaling molecules, but it can't, e.g., aggregate vast numbers of visual neurons through multiple layers of neural networks in order to identify a high-level object like "angry person with an axe in his hand". Complex internal representations are required for high degrees of intelligence. In other words, representations aren't relevant in themselves but only because of the cognitive possibilities they open up.
Of course, unicellular organisms have their own sets of cognitive possibilities, and different environmental situations (i.e., different "mental representations" in the extended mind) open up different cognitive possibilities. For example, in a similar way as a person's mental representation of an axe-wielding man will yield different thoughts/behaviors than a mental representation of a cute bunny, so too the environmental context for a bacterium in the gut triggers different "thoughts"/behaviors by the bacterium than the environmental context of a hydrothermal vent triggers in a bacterium living there.
My point here is not that bacteria are equivalent to more intelligent organisms. Rather, my aim (in this and many of my writings on consciousness) is to problematize the intuitive distinctions that we often make when trying to separate minds from non-minds.
Favela (2017): "organisms do not need to perceive the world indirectly via mental images/representations in order to have phenomenal consciousness. [...] If the world serves as its own best model (Brooks, 1991), then environments can serve as rich sources of information. If so, then there would be no need to appeal to representations to explain how organisms conduct successful goal-directed and adaptable behavior" (pp. 1-2).
Valence
Lyon (2015):
Valence refers to the attractiveness, acceptability or tolerability of a stimulus [...].
[In bacteria,] the valence of a signal is implicit in the processes that coordinate, say, the rotation bias of flagellar motors or the complex developmental sequence of signaling, genetic transcription and protein expression that leads to sporulation. [...]
Virtually nothing is known, in bacteria or humans, about how combinations of external stimuli of differing valence are integrated with interoceptive cues into a coherent behavioral response.
Comparison with neural networks
Fernando et al. (2008):
recent work in systems biology reveals molecular circuits that are rather similar to neural networks (Bray 2003) and logic gates (Buchler et al. 2003) within individual cells. [...] chemical kinetics is Turing universal (Magnasco 1997) and can therefore at least in principle implement arbitrary neural networks (Hjelmfelt et al. 1991) and finite state automata (Hjelmfelt et al. 1992)[...].
Freddolino and Tavazoie (2012), p. 377:
Regulatory networks in microbes and neural networks in metazoa have essentially the same function: to integrate all sensory information available to the organism, estimate on the basis of that information (and prior knowledge, genetic and acquired) the likely current and future states of the organism’s environment, and optimize the organism’s behavior in light of those predictions. The importance of integration of available information and subsequent prediction of environmental states has been appreciated in the context of animal behavior for decades [...], but the presence of similarly complex analysis and predictive behavior in microbes has been discovered only recently [...].
Computation by single neurons
While neurons of animals are not "unicellular organisms" in the usual sense, they are single cells, and it's interesting to learn about their own individual "cognitive abilities".
It turns out that a single neuron of a multicellular animal is a fairly complex computing device in its own right. One example of this point can be seen from the field of "dendritic computing".
Senior author Professor Michael Hausser commented: “This work shows that dendrites, long thought to simply ‘funnel’ incoming signals towards the soma, instead play a key role in sorting and interpreting the enormous barrage of inputs received by the neuron. Dendrites thus act as miniature computing devices for detecting and amplifying specific types of input.["]
London and Häusser (2005) review the topic of dendritic computation. Here's one tidbit from the paper (p. 512):
Each subunit thus integrates its inputs and passes them through a [sigmoidal] nonlinearity function. This gives each piece of dendrite the computational power of a small unit similar to those conventionally used in neural networks. The output of each subunit is conveyed to the soma, in terms of passive dendritic integration. The picture that emerges from this analysis is of a two-layered neuronal network that resides within a single neuron. This analysis is supported by detailed modeling of single neurons, showing that the predictive power of the two-layer neural network description is very good [...], and by experiments in layer 5 pyramidal neurons [...]. The attractiveness of this approach stems from the fact that two-layer neural networks are general-purpose computation machines, which have been extensively studied and can implement very powerful computations.
Footnotes
- However, strangely, Brady (1974) says something different on p. 115: "It is estimated that 60-80 percent of the total soil metabolism is due to the microflora. Not only do they destroy plant residues but they function in the digestive tracts of animals and eventually decompose the dead bodies of all organisms." (back)